3 tryby transferu genetycznego w komórkach bakterii
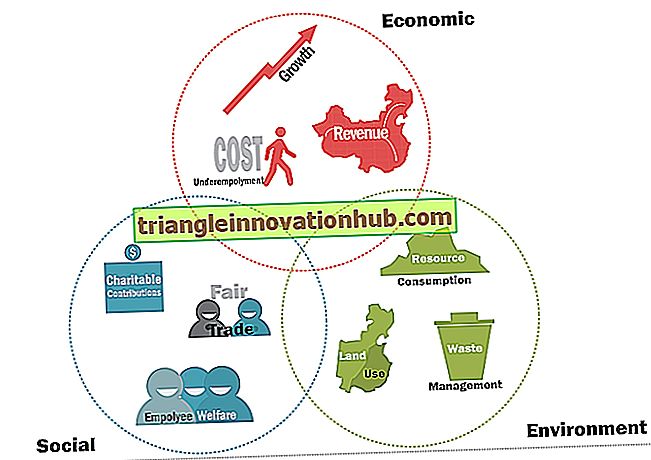
Trzy tryby transferu genetycznego w komórkach bakteryjnych to: (a) transformacja, (b) transdukcja, (c) koniugacja.
Bakterie dzielą się bardzo szybko. Czas podwojenia nazywany jest również czasem generacji i może wynosić zaledwie 20 minut. Bakterie rozmnażają się głównie poprzez rozmnażanie bezpłciowe, ale nie wykazują prawdziwej reprodukcji płciowej, ponieważ nie wytwarzają fazy diploidalnej. Tak więc brakuje mejozy. Jednak bakterie wymieniają materiał genetyczny między dwiema komórkami.
Sposoby transferu genetycznego bakterii:
Trzy tryby przenoszenia genetycznego między komórkami bakteryjnymi to:
(a) Transformacja
(b) Transdukcja
(c) Koniugacja.
(a) Transformacja:
Zjawisko polegające na tym, że DNA wyizolowany z jednego typu komórki, gdy jest wprowadzony do innego typu, może nadać niektóre swoje właściwości temu drugiemu, nazywa się transformacją. Zostało to potwierdzone przez Griffitha w jego eksperymentach na bakteriach Streptococcus pneumonia.
(b) Transdukcja:
Transfer materiału genetycznego z jednej bakterii do drugiej przez bakteriofag jest nazywany transdukcją.
(c) Koniugacja:
Jednokierunkowy transfer DNA z jednej komórki do drugiej przez mostek cytoplazmatyczny nazywany jest koniugacją. Proces ten jest równoznaczny z kojarzeniem seksualnym u eukariontów. Dwie bakteryjne komórki haploidalne różnych szczepów zbliżają się do siebie.
Poznają się wzajemnie poprzez komplementarne makrocząsteczki na ich powierzchni. Dawca lub komórka męska przekazuje część lub całość chromosomu do komórki biorcy lub komórki żeńskiej. Zdolność przenoszenia materiału genetycznego od samca kontrolowana jest przez czynnik płeć lub płodność (gen F) obecny w plazmidzie.
Tak więc, geny mogą być przenoszone z dawcy do komórki biorcy na cząsteczce DNA, która działa jako czynnik płciowy zwany genem F. Ten gen płciowy może znajdować się w chromosomie bakteryjnym lub może istnieć jako autonomiczna jednostka w cytoplazmie.
Bakterie męskie z ciernistymi wypukłościami zwanymi piliami płciowymi wchodzą w kontakt z żeńską bakterią, która nie ma pilusa i nie przekazuje swojego DNA. Czynnik F (plazmid) przenosi geny odpowiedzialne za produkcję pili i inne funkcje wymagane do przeniesienia DNA. Czasami współczynnik F integruje się z chromosomem bakteryjnym.
Takie bakterie mogą przenosić swój materiał genetyczny do żeńskiej komórki z wysoką częstotliwością (Hfr) w określonej sekwencji. Nazywa się je Hfr -strains. Koniugację po raz pierwszy wykazali Lederberg i Tatum w E. coli. Częstotliwość rekombinacji była bardzo niska w eksperymentach Lederberga.
Komórka Hfr działa jak męska bakteria i po zmieszaniu z żeńską (F-) komórką tworzy most koniugacyjny. Czynnik F zawierający DNA pęka w określonym punkcie i rozpoczyna wstawianie DNA do żeńskiej, a sekwencja przenoszenia genu chromosomalnego jest zawsze w tej samej kolejności (geny A, B, C i D).
Współczynnik F jest przenoszony na końcu. Mostek koniugacji zazwyczaj pęka zanim przeniesiony zostanie cały chromosom. Tylko geny A i B zostały przeniesione w podanym przykładzie. Te geny A i / lub B mogą rekombinować z odpowiednimi genami w chromosomie F.
Tak więc, jeśli B 'w komórkach F jest zmutowaną postacią B, kradzież B' w chromosomie F może stać się B w wyniku rekombinacji po koniugacji. Zatem markery genetyczne można przenosić z gospodarza do odpowiedniego odbiorcy bez takich markerów.
Kolejność, w jakiej takie markery są przekazywane do biorcy, byłaby zgodna z kolejnością, w jakiej są obecne w dawcy. Zatem eksperymenty koniugacji są użyteczne w konstruowaniu map genów (kolejności rozmieszczenia genów w chromosomie) organizmów.
Hayes (1952) znalazł szczep E. coli, w którym częstotliwość rekombinacji była tak wysoka jak 100 do 1000 razy, jak donosi Lederberg. Szczep nazwano szczepem rekombinowanym (Hfr) o wysokiej częstotliwości.